

Faeq M. Zaman 1*, Jon A. Friesen1
1 Illinois State University
ABSTRACT
Wheat germ acid phosphatase (WGAP) is a commonly used protein in laboratory settings for studying the release of inorganic phosphate (Pi) from phosphate esters under acidic conditions. When kinetically quantifying the enzyme, the typical procedure involves a spectrophotometric assay that indirectly measures WGAP activity. In this study, a new assay was developed to directly measure WGAP activity, involving the integration of a 31P NMR resonance peak representing the Pi product with a chemical shift of δ = 5.25 ppm. In order to compare the effectiveness of this new assay with the traditional method, both the 31P NMR spectroscopy-based assay and the traditional spectrophotometric-based assay were used side-by-side to evaluate WGAP activity. The extent to which activity was measured accurately using a UV-Visible spectrophotometer was quite limited, as the instrument only allowed accurate absorbance readings at relatively lower concentrations. Alternatively, performing analysis with 31P NMR spectroscopy introduced many advantages as the isotope is naturally abundant and the detection of the 31P signal at higher concentrations was possible. The total average WGAP specific activity determined from the spectrophotometric assays was 0.436 ± 0.023 μmol product/min/mg enzyme, and for the 31P NMR assays was 0.368 ± 0.098 μmol product/min/mg enzyme. When the two datasets of specific activity values from each method were compared, the analysis revealed a statistically significant similarity between the medians, thus confirming the viability of the new assay. These initial studies indicate that an assay based upon the integration of 31P NMR spectroscopy could be used for kinetic analysis of any reaction involving phosphate transfer.
Keywords: 31P NMR, NMR spectroscopy, enzyme assay, enzyme kinetics, acid phosphatase
AUTHOR SUMMARY
Performing enzyme assays is a fundamental skill that students in our lab learn as they are introduced to the subject of biochemistry. In the typical undergraduate biochemistry curriculum, students usually only learn one method to evaluate enzyme kinetics, that is, spectrophotometrically. Little do they realize that catalytic activity can be examined through a diverse range of methods. Therefore, to familiarize our students with advanced chemical equipment and alternative enzyme analysis techniques, we attempted to design and implement a novel enzyme activity assay experiment using 31P NMR spectroscopy. We wanted to explore how using a different technique might impact the ability to evaluate enzyme activity accurately and consistently. By demonstrating the validity of using 31P NMR to measure acid phosphatase activity, we believe our project contributes to the expansion of the applications of NMR spectroscopy beyond structural determination.
INTRODUCTION
Acid phosphatase from wheat germ (WGAP) is a non-specific enzyme that catalyzes the hydrolysis of phosphate monoesters (Kalita & Ambasht, 2019). Although not firmly established, WGAP is presumed to have a role in the mobilization and utilization of phosphate reserves of the seed for plant nutrition (Hasan et al., 2016). Extracted from the embryo of wheat plants, WGAP is a commercially prepared protein that is often used in biochemistry laboratory settings to catalyze dephosphorylation reactions (Moorman & Brayton, 2021). The enzyme best operates in an acidic environment, hence its classification as an acid phosphatase (Kalita & Ambasht, 2019). One substrate WGAP can hydrolyze is p-nitrophenyl phosphate (pNPP), producing p-nitrophenol (pNP) and inorganic phosphate (Pi) as products (Figure 1). When the pNP product is deliberately deprotonated, it forms the anionic form of pNP, known as p-nitrophenolate (pNP-), which conveniently absorbs light within the visible spectrum at a wavelength maximum of 405 nm (Malathi et al., 2014). Consequently, the quantification of pNP and the evaluation of WGAP activity using a spectrophotometer become feasible.
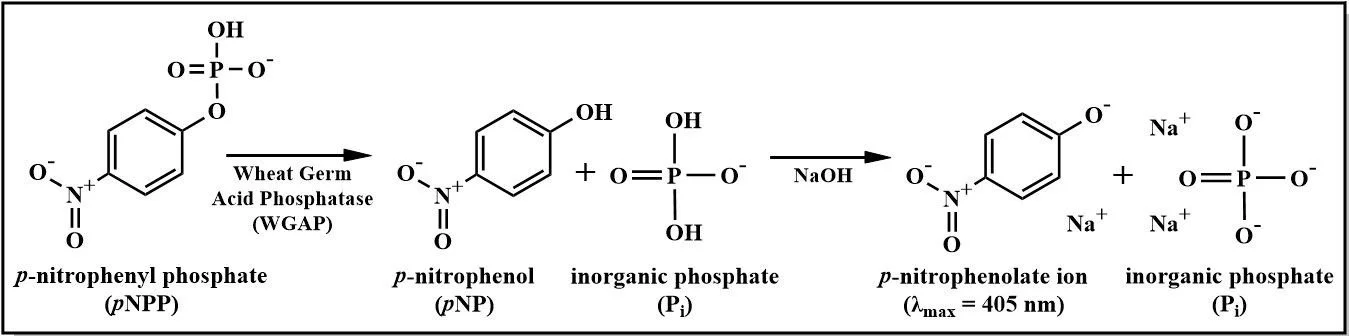
Figure 1. Hydrolysis of p-Nitrophenyl Phosphate (pNPP) Reaction Diagram. The reaction shown is catalyzed by the enzyme under study, that is, by acid phosphatase derived from wheat germ. Subsequent deprotonation of the products allowed for spectrophotometric quantification of pNP-, and therefore quantification of the pNP product, at 405 nm. The other product, Pi, was quantified using 31P NMR spectroscopy by integrating the associated resonance peak.
The typical spectrophotometric method of measuring WGAP activity is dependent on both the extra step of adding base to deprotonate the product as well as the use of the Beer-Lambert Law to convert absorbance values to concentration. By applying acid-base equilibria principles and utilizing the Henderson-Hasselbach equation, the addition of excess base causes a significant shift in the equilibrium, resulting in the pNP concentration becoming negligible compared to pNP-. As a result, the concentration of pNP- that is measured is essentially equal to the concentration of pNP product produced by WGAP prior to adding base. Despite its simplicity and reliability as an assay method, the inclusion of additional steps that lead to indirect measurements of WGAP activity is a cause for concern due to the potential for reduced precision. Furthermore, the typical UV-Visible spectrophotometers used in these assays present limitations as their highest sensitivity and accuracy lie in detecting compounds at low concentrations, which restricts the linearity of the absorbance-concentration relationship (L.C. Passos & M.F.S. Saraiva, 2019).
To address these limitations, an alternative technique utilizing phosphorus-31 nuclear magnetic resonance (31P NMR) spectroscopy was explored in this study. NMR spectroscopy is a powerful analytical tool that exploits the magnetic properties of atomic nuclei. By subjecting the sample to a strong magnetic field and radiofrequency pulses, the nuclei in the sample absorb and emit electromagnetic radiation at specific frequencies, generating a unique NMR spectrum. This spectrum provides valuable information about the elements present and the arrangement of atoms in molecules, which is crucial for monitoring chemical reactions (Martin, 2020). In the case of 31P NMR spectroscopy, it specifically allows for the detection, relative quantification, and analysis of phosphorus-containing compounds (Stücker et al., 2018). This technique offers several distinct advantages, such as its ability to detect a wide range of concentrations, as well as its utilization of the naturally abundant phosphorus-31 isotope (Walker et al., 2019; Jarosch et al., 2015). These properties make 31P NMR well-suited for directly measuring the formation of the other product produced in the WGAP reaction, inorganic phosphate (Pi), and evaluating WGAP activity.
Understanding the activity of WGAP and its hydrolysis of phosphate monoesters is highly relevant in the field of agriculture, particularly in crop production. Acid phosphatases, including WGAP, serve as indicators of soil quality, highlighting their importance in assessing soil health (Attademo et al., 2021). Access to phosphorus is crucial for optimal plant growth, but often, phosphorus becomes inaccessible to roots due to its insoluble complexes with metal cations or fixations as organophosphate in the soil (Bhadouria & Giri, 2021; Richardson & Simpson, 2011; Dissanayaka et al., 2018). As plants can only absorb and utilize soluble inorganic phosphates (Pi), hydrolases like acid phosphatases play a vital role in releasing soluble Pi and maintaining plant homeostasis, particularly under Pi-deficient conditions (Upadhyay et al., 2022). Expanding our knowledge of WGAP’s kinetic characteristics can therefore assist scientists in developing targeted strategies for improving nutrient uptake and utilization in plants, leading to enhanced agricultural sustainability and increased crop yield (Dotaniya et al., 2019; Hasan et al., 2016). Additionally, the development of advanced techniques for directly measuring WGAP activity provides researchers with a more accurate and precise means of evaluation, while also broadening the range of analytical tools for studying enzymatic reactions. Therefore, by incorporating 31P NMR spectroscopy, this research aims to contribute to a deeper understanding of enzyme kinetics. Additionally, it particularly provides valuable insights into the catalytic properties of WGAP and further advances our understanding of its role in agricultural contexts.
MATERIALS AND METHODS
Materials and Instrumentation
Sodium acetate (NaCH3COO), acetic acid (CH3COOH), monosodium phosphate (NaH2PO4), and sodium hydroxide (NaOH) were obtained from Fisher Scientific. Wheat germ acid phosphatase (WGAP), p-nitrophenyl phosphate (pNPP), p-nitrophenol (pNP), and magnesium chloride (MgCl2) were sourced from Sigma Aldrich. Deuterium oxide (D2O) was acquired from Cambridge Isotope Laboratories. Using NaCH3COO and CH3COOH, a stock solution of 0.5 M acetate buffer at pH 4.5 was prepared. This buffer was used for all sample preparations and experiments in this study. Using this buffer, stock solutions of WGAP (1 mg/mL) and pNPP (5 mg/mL) were also made fresh. The spectrophotometric assays were conducted using a Thermo Scientific Evolution 260 Bio UV-Visible spectrophotometer. For assays designated for 31P NMR analysis, a 400 MHz Bruker Avance III NMR spectrometer with a broadband probe was utilized. The following instrument parameters were employed for NMR data collection: total data points (TD) = 16384, data size (DS) = 4, number of scans (NS) = 300, spectral width (SW) = 50 ppm, relaxation delay frequency (RO) = 20 Hz, relaxation delay time (D1) = 2 sec, and the receiver gain = 1030. NMR spectra were analyzed using Bruker TopSpin 3.6.5 software.
Determination of the pNP- Molar Absorptivity Constant
To determine the molar extinction coefficient of pNP-, a series of ten solutions with increasing concentrations of pNP (ranging from 0.01 mM to 0.1 mM) were prepared. Sodium hydroxide (NaOH) was added to achieve a final concentration of 500 mM, converting pNP to pNP-. Using a 1 cm path length cuvette, the absorbance of each solution was measured at 405 nm and a plot was constructed to correlate the absorbance values with the known pNP- concentrations. The molar absorptivity constant of pNP- at this wavelength was obtained by calculating the slope of the linear regression plot. This constant was subsequently used to determine the pNP product concentrations in the later spectrophotometric WGAP assays.
Substrate and Product Standard 31P NMR Chemical Shifts
To ensure minimal overlap in the phosphorus resonances of the substrate (pNPP) and product (Pi), the 31P NMR chemical shifts for these compounds were determined. A standard mixture containing 5 mM pNPP, 5 mM NaH2PO4, 5 mM MgCl2, and 500 mM NaOH in buffer was prepared and analyzed. The NMR spectrum of this standard mixture allowed for the identification and distinction of the 31P peaks of the substrate and product, which was critical for the accurate integration and determination of their respective concentrations as WGAP releases Pi from pNPP.
Wheat Germ Acid Phosphatase Enzyme Assays
In this investigation, two sets of assays were performed and compared. The first set involved measuring the absorbance of pNP- at 405 nm, while the second set was based on the new method of integrating the 31P NMR signal of deprotonated Pi. Experiments for both techniques consisted of one control assay and four experimental assays performed in triplicate, resulting in a total of 13 assays (12 experimental assays and 1 control assay) for each method. For both sets of assays, two experiments were performed: an enzyme-dependent experiment and a time-dependent experiment. All assay tubes had identical conditions: total volume = 1 mL, final WGAP metal cofactor concentration = 5 mM MgCl2, incubation temperature = 37ºC water bath, and pH = 4.5. To ensure that substrate concentrations remained higher than enzyme concentrations in every assay, the pNPP concentration was held constant at 3.368 mM in all assays prior to their reactions (using 250 µL of the stock solution in each assay). Each reaction was initiated by adding pNPP and terminated by adding 50 μL of 10 M NaOH.
After determining what combinations of enzyme concentrations and reaction times resulted in product concentrations falling within the detection limits of the respective instruments, the enzyme-dependent experiment for both sets of assays was conducted with varying amounts of WGAP and 30 minutes of incubation time. The enzyme concentrations used for the spectrophotometric assays ranged from 0.043 μM to 0.215 μM, while the enzyme concentrations used for the 31P NMR method ranged from 0.258 μM to 1.204 μM. Initially, we attempted to conduct a single set of assays that could be split afterwards and analyzed on both instruments simultaneously. However, we encountered differences in the detection ranges between the instruments, with several assays producing small Pi 31P NMR signals but high absorbance values of pNP-. This is why separate assays, of differing enzyme concentration ranges, were performed for the two methods. After stopping the reactions, the first set of assays was promptly analyzed for absorbance at 405 nm. For the second set of assays, 500 μL of each assay mixture was transferred to a new microcentrifuge tube, and 55 μL of D2O was added to obtain a final 90:10 H2O:D2O ratio for NMR lock. These solutions were then pipetted into 5 mm NMR tubes to collect 162 MHz 31P NMR data.
Following the analysis of the enzyme-dependent experiment, a suitable WGAP concentration within the first-order rate was selected for each assay method for use in the subsequent time-dependent experiment. In this experiment, for both assay methods, the enzyme concentration remained constant while the incubation time varied. The reaction mixtures designated for spectrophotometric analysis contained 0.129 µM WGAP, while those designated for 31P NMR analysis contained 1.204 µM WGAP. Data for each method were collected in the same manner as done for the enzyme-dependent experiment. When performing analyses via the spectrophotometric method, the no-enzyme assay and the zero-time assay served as controls for the experiments. These control assays were then used to blank the spectrophotometer before reading the results of the experimental assays.
Measuring Product Formation Using the Two Assay Methods
For all enzyme assays analyzed via the traditional spectrophotometric method, the Beer-Lambert Law equation (A405nm = εbc) was used to convert the measured 405 nm absorbance values to pNP- concentrations. In the Beer-Lambert Law equation, A represents the absorbance at 405 nm, ε is the molar absorptivity constant determined from the first experiment in this research, b is the path length of 1 cm, and c is the concentration of pNP-. As explained in detail previously, the concentrations calculated in this step were also equal to the concentrations of pNP produced by WGAP. To calculate the Pi concentrations from enzyme assays designed for 31P NMR analysis, the control sample in each experiment, specifically the no-enzyme sample in the enzyme-dependent experiment and the zero-time sample in the time-dependent experiment, were first integrated. Since all control and experimental samples initially contained 3.368 mM pNPP, the corresponding 31P signal integration of pNPP for the control samples were set as the baseline from which all other peaks and their respective concentrations were calculated. For example, when the integration of a control substrate peak at δ = -0.34 ppm yielded a value of 2.67×107 and the integration of an experimental product (Pi) peak at δ = 5.25 ppm yielded a value of 1.50×106, the concentration of Pi was calculated to be 0.1892 mM using a simple proportion.
Comparing the New 31P NMR-Based Assay with the Spectrophotometric-Based Assay
To assess the validity and effectiveness of the newly developed assay method, the results obtained from the 31P NMR-based assays were compared directly with the results obtained from the conventional spectrophotometric assays. Given that the enzyme concentrations differed between the two sets of assays, the most appropriate approach for comparison was to calculate the specific activities of the assays from each method. Specific activity normalizes the rate of product formation per milligram of enzyme used in the assay (Walker et al., 2019). Therefore, specific activity values were expressed in units of μmol product formed per minute per mg of WGAP. It is worth noting that specific activity can be influenced by varying substrate concentrations. However, in this study, we maintained consistent substrate concentrations between the spectrophotometric assays and 31P NMR assays. With equal initial substrate concentrations, the two assay methods were therefore expected to yield similar specific activity values for WGAP. This allowed for a direct comparison between the 31P NMR-based assay and the traditional spectrophotometric assay.
Calculating the specific activities of the reactions in this study involved several steps. The amount of product formed in each reaction, measured in μmol, was determined by multiplying the final product concentration, obtained either through the Beer-Lambert Law or through integrating a 31P NMR resonance peak, with the total assay volume. Dilutions made during reaction termination and NMR sample preparation were accounted for in the calculations. The amount of enzyme used in each reaction, measured in milligrams, was determined by considering the dilution that occurred each time a volume from the 1 mg/mL WGAP stock solution was added to an assay tube. Once the micromoles of product and milligrams of WGAP were determined, the specific activity value was obtained by dividing the amount of product formed by the reaction time and mass of enzyme used.
To compare the two assay methods, the total average specific activity value for each method was calculated by pooling the data from both the enzyme-dependent experiment and the time-dependent experiment. To determine whether the medians of the two datasets were similar, a two-tailed Mann-Whitney U test was performed at the α = 0.05 significance level. This particular statistical test was chosen because the two assay groups were independent, not assumed to follow a normal distribution, nor assumed to have similar variances between each other (Usman, 2016).
RESULTS
Establishing the pNP- Molar Extinction Coefficient at 405 nm
From the experiment involving the preparation of ten different pNP solutions, their deprotonation, and subsequent analysis on the spectrophotometer, the plot that was constructed revealed a clear linear relationship between absorbance at 405 nm and pNP- concentrations (Figure 2). The linear regression analysis of the plot resulted in a slope value of 16,500 M-1cm-1, which corresponds to the pNP- molar absorptivity constant at this specific wavelength. This value was crucial for the application of the Beer-Lambert Law equation in calculating the pNP product concentrations in the spectrophotometric-based assays.

Figure 2. The Effect of p-Nitrophenolate Ion Concentration on Absorbance at 405 nm. To determine the molar absorptivity constant of pNP-, absorbance at the wavelength maximum of pNP- (405 nm) was measured at increasing concentrations of pNP- using a 1 cm path length cuvette and a UV-Visible spectrophotometer. Using the Beer-Lambert Law, the slope of the regression line shown gives the molar absorptivity of pNP-. This constant (16,500 M-1cm-1) was then used to quantify pNP production in the enzyme assays.
31P NMR Chemical Shifts of Deprotonated pNPP and Pi
In order for the use of 31P NMR spectroscopy to be a reliable method for measuring the catalytic activity of WGAP, it is essential that the 31P NMR resonances associated with the substrate (pNPP) and product (Pi) do not overlap in the spectrum. When the standard solution containing deprotonated pNPP and Pi was subjected to 31P NMR analysis, clear separation of peaks was observed in the resulting NMR spectrum. The peak corresponding to pNPP was observed at δ = -0.34 ppm, while the peak corresponding to Pi was observed at δ = 5.25 ppm (Figure 3). The distinct separation of these peaks allowed for the integration of peak areas and their correlation with the concentrations of the substrate and product.

Figure 3. 162 MHz 31P NMR Spectrum of Standard Sample. A standard sample containing a mixture of pNPP and Pi in acetate buffer pH 4.5 was created. The 31P NMR spectrum of this sample was obtained to determine the chemical shifts of the pNPP substrate (-0.34 ppm) and Pi product (5.25 ppm) that would be present in the WGAP catalyzed reactions. During the enzyme assays, Pi formation was quantified by integrating the associated 31P resonance peak.
Analyzing the Activity of WGAP Using Spectrophotometry
The activity of WGAP in producing pNP through the hydrolysis of pNPP was first assessed by varying the enzyme concentration. In these spectrophotometric assays, a linear relationship was observed between WGAP concentration and pNP production, indicating that increasing the enzyme concentration resulted in higher levels of pNP production (Figure 4). As expected, the no-enzyme condition exhibited no detectable absorbance, confirming that pNP production did not occur in the absence of WGAP. This supported the use of the no-enzyme assay mixture as the spectrophotometric blank for the enzyme-dependent experiment. In the following time-dependent experiment, WGAP activity was measured as a function of incubation time while keeping the enzyme concentration constant at 0.129 µM. The effect of reaction time on WGAP’s production of pNP in these spectrophotometric assays displayed a strong positive linear relationship (Figure 5). Similar to the no-enzyme assay, the zero-time assay demonstrated negligible absorbance, indicating the absence of product formation at the start of the reaction. Therefore, the use of the zero-time assay mixture as the spectrophotometric blank for the time-dependent experiment was appropriate.

Figure 4. pNP Formation as a Function of WGAP Concentration Using the Spectrophotometric Method. The plot produced here depicts an apparent linear relationship between pNP formation and increasing WGAP concentration. The WGAP concentration of 0.129 µM, which was within the range of concentrations seen in this plot, was selected for the subsequent time-dependent experiment.

Figure 5. pNP Formation as a Function of Reaction Time Using the Spectrophotometric Method. The graph shown depicts an apparent first-order relationship between pNP formation and reaction time. Dividing the resulting reaction rates by the amount of enzyme used allowed for the determination of WGAP specific activity.
Analyzing the Activity of WGAP Using 31P NMR Spectroscopy
Following a similar approach to the spectrophotometric method, the activity of WGAP in hydrolyzing pNPP and releasing Pi was analyzed using 31P NMR spectroscopy. The plot generated from the 31P NMR assays in the enzyme-dependent experiment, correlating Pi formation with WGAP concentration, revealed a clear positive linear relationship (Figure 6). Consistently, the no-enzyme assays utilizing the 31P NMR method displayed no detectable Pi 31P resonance signal in the NMR spectra, indicating the absence of product production without the presence of WGAP. For the subsequent time-dependent experiment, where the WGAP concentration in the 31P NMR assays was kept constant at 1.204 μM WGAP, the data obtained demonstrated a linear increase in Pi formation with increasing reaction time (Figure 7). As expected, the zero-time assay showed no observable Pi 31P resonance signal, confirming that no product formation occurs without sufficient time for WGAP catalysis.

Figure 6. Pi Formation as a Function of WGAP Concentration Using the 31P NMR-Based Method. The plot produced here depicts an apparent positive linear relationship between Pi formation and increasing WGAP concentration. The WGAP concentration of 1.204 µM, which was within the range of concentrations seen in this plot, was selected for the subsequent time dependent experiment.

Figure 7. Pi Formation as a Function of Reaction Time Using the 31P NMR-Based Method. The graph shown depicts an apparent first-order relationship between Pi formation and reaction time. Dividing the resulting reaction rates by the amount of enzyme present in the assays allowed for the determination of each assay’s WGAP specific activity. These specific activity values could then be compared to the specific activity values obtained from the spectrophotometric method.
WGAP Specific Activity Values from Each Assay Method and Significance Test Results
When applying the spectrophotometric method, the average of the specific activity values calculated from the enzyme-dependent experiment was 0.425 ± 0.009 μmol pNP/min/mg WGAP, while the average from the time-dependent experiment was 0.448 ± 0.030 μmol pNP/min/mg WGAP. When employing the 31P NMR spectroscopy method, the average of the specific activity values obtained from varying enzyme concentration was 0.300 ± 0.018 μmol Pi/min/mg WGAP, while the average obtained from varying reaction time was 0.436 ± 0.099 μmol Pi/min/mg WGAP. When looking at the spread of specific activity values for each assay method, the interquartile ranges of the two methods do indeed overlap; however, it is visibly clear that the values obtained from the 31P NMR assays were much more dispersed than those obtained from the spectrophotometric method (Figure 8). Comparing the overall average specific activity between the two techniques, the results were quite similar. The overall average WGAP specific activity from all the spectrophotometric assays was 0.436 ± 0.023 μmol product/min/mg WGAP, while from all the 31P NMR assays was 0.368 ± 0.098 μmol product/min/mg WGAP. Conducting a Mann-Whitney U test on the specific activity values from each assay method yielded a U-value of 15, a z-score of 1.733, and a p-value of 0.084. Since the calculated U-value was greater than the critical U-value at the 0.05 alpha level (13) and the p-value was above 0.05, the difference between the medians was not found to be statistically significant.

Figure 8. Project Data Summary: Comparing the Specific Activity Values Between the Spectrophotometric Assays and the 31P NMR Assays. To establish the validity of the 31P NMR-based assay to kinetically characterize WGAP, the average specific activity values from all assays performed using each assay method was compared. Although the spread of the specific activity values from the 31P NMR method are visibly much more dispersed than those from the spectrophotometric method, performing a Mann-Whitney U test revealed that the medians of the two specific activity spreads were not significantly different at the 0.05 significance level (U = 15, z-score = 1.733, and p > 0.05). This suggested that the 31P NMR method can measure WGAP activity just as well as the traditional spectrophotometric method.
DISCUSSION
In this study, we determined the molar extinction coefficient at 405 nm for pNP- to be 16,500 M-1cm-1, which is in exact agreement with what is reported in existing literature (Chaudhuri et al., 2013). To assess WGAP activity, we employed two distinct assay methods: the traditional absorbance-based assays and the novel 31P NMR-based assays. While both methods aimed to measure the concentration of a product molecule resulting from WGAP catalysis, they differed in their approaches. Specifically, the spectrophotometric assays indirectly measured pNP production by detecting its deprotonated form, while the 31P NMR-based assays allowed for direct measurements of Pi formation. By demonstrating the capability of the 31P NMR technique to directly measure product formation, our study sought to establish its validity as a reliable alternative to the traditional spectrophotometric method for assessing enzyme activity, such as in the case of WGAP.
To optimize the enzyme concentrations used in each assay, it was crucial to ensure that the amount of product formed in each assay fell within the detection limits of the respective instruments. The linear relationships observed in our experiments confirmed that we had indeed selected suitable enzyme concentrations for each method; the reactions had proceeded successfully, and the resulting product signals were not too weak or too strong for the instruments to make reliable measurements. These observations have therefore highlighted the different concentration ranges that each assay method is suited for, with the spectrophotometric assay being tailored towards analyzing samples of lower concentrations and the 31P NMR assay being better suited for samples of higher concentrations (L.C. Passos & M.F.S. Saraiva, 2019).
The relatively large dispersion observed in the specific activity values obtained from the 31P NMR method could be attributed to a few factors. Firstly, when dealing with assays with incubation times of less than 5 minutes, we encountered difficulties in perfectly managing all the assays simultaneously. Slight delays in starting and stopping these reactions may have occurred, introducing inaccuracies and inconsistencies in the results. These possible delays could have led to slightly higher specific activity values than expected. In future studies, we recommend using longer incubation times that are more manageable and easier to work with. Additionally, increasing the number of replicates and number of assays for each experiment could enhance accuracy and precision. Therefore, performing at least five replicates per assay and conducting at least five different experimental assays per experiment for each method would help improve data variance and enhance accuracy.
Despite the differences in dispersion between the two methods, the results of the Mann-Whitney U test indicated that the medians of the specific activity values obtained from each method were not significantly different. This finding strongly suggests that measuring WGAP activity using 31P NMR spectroscopy yields similar results to the spectrophotometric method. Therefore, 31P NMR spectroscopy can be considered a viable and effective technique for characterizing the hydrolysis of pNPP by WGAP. The development of this new assay method, which allowed the direct calculation of reaction rates from the reaction products, was a pivotal advancement. While qualitative studies using 31P NMR to investigate WGAP have been conducted previously (Van Etten & Waymack, 1991), a distinct method for kinetically evaluating WGAP using 31P NMR had not yet been established. By pioneering this work, we not only enabled the quantification of Pi formation but also introduced the utilization of an instrument with a much wider detection range compared to the traditional spectrophotometer.
The application of 31P NMR spectroscopy in detecting phosphate-containing products in enzyme-catalyzed reactions demonstrated in this study holds great potential for investigating other enzymes involving phosphate-containing substrates or products. The direct measurement of phosphate-containing products using techniques like 31P NMR spectroscopy can provide significant insights into the catalytic activity and kinetics of these enzymes (Walker et al., 2019; Jarosch et al., 2015). Furthermore, the WGAP activity values obtained in this study can serve as a valuable resource for agricultural researchers, contributing to the understanding and management of soluble Pi availability in soils and its uptake in plants (Dotaniya et al., 2019; Hasan et al., 2016).
As a whole, this study successfully compared the newly developed 31P NMR-based assay with the traditional spectrophotometric assay to measure WGAP activity, validating the effectiveness of the new method. The findings underscore the value of 31P NMR spectroscopy in characterizing enzymatic reactions and emphasize its wide range of potential applications.
REFERENCES
Attademo, A.M., Sanchez-Hernandez, J.C., Lajmanovich, R.C., Repetti, M.R. and Peltzer, P.M. (2021). Enzyme Activities as Indicators of Soil Quality: Response to Intensive Soybean and Rice Crops. Water, Air, & Soil Pollution, 232(7). doi:10.1007/s11270-021-05211-2
Bhadouria, J. and Giri, J. (2021). Purple acid phosphatases: roles in phosphate utilization and new emerging functions. Plant Cell Reports, 41(1), 33–51. doi:10.1007/s00299-021-02773-7
Chaudhuri, G., Chatterjee, S., Venu-Babu, P., Ramasamy, K., & Thilagaraj, W. R. (2013). Kinetic behaviour of calf intestinal alkaline phosphatase with pNPP. Indian Journal of Biochemistry & Biophysics, 50(1), 64–71. http://nopr.niscpr.res.in/handle/123456789/16064
Dissanayaka, D.M.S.B., Plaxton, W.C., Lambers, H., Siebers, M., Marambe, B. and Wasaki, J. (2018). Molecular mechanisms underpinning phosphorus-use efficiency in rice. Plant, Cell & Environment, 41(7), 1483–1496. doi:10.1111/pce.13191
Dotaniya, M.L., Aparna, K., Dotaniya, C.K., Singh, M. and Regar, K.L. (2019). Role of Soil Enzymes in Sustainable Crop Production. Enzymes in Food Biotechnology, 569–589. doi:10.1016/b978-0-12-813280-7.00033-5
Hasan, Md. M., Hasan, Md. M., Teixeira da Silva, J. A., & Li, X. (2016). Regulation of phosphorus uptake and utilization: transitioning from current knowledge to practical strategies. Cellular & Molecular Biology Letters, 21(1). doi:10.1186/s11658-016-0008-y
Jarosch, K. A., Doolette, A. L., Smernik, R. J., Tamburini, F., Frossard, E., & Bünemann, E. K. (2015). Characterisation of soil organic phosphorus in NaOH-EDTA extracts: A comparison of 31P NMR spectroscopy and enzyme addition assays. Soil Biology and Biochemistry, 91, 298–309. doi:10.1016/j.soilbio.2015.09.010
Kalita, T., & Ambasht, P. K. (2019). Immobilization and characterization of acid phosphatase from wheat germ (Type I) in Agarose gel. Journal of Proteins and Proteomics, 10(4), 291– 297. doi:10.1007/s42485-019-00023-9
L.C. Passos, M. and M.F.S. Saraiva, M.L. (2019). Detection in UV-visible spectrophotometry: Detectors, detection systems, and detection strategies. Measurement, 135, 896–904. doi:10.1016/j.measurement.2018.12.045
Malathi, S., Ezhilarasu, T., Abiraman, T., Balasubramanian, S. (2014). One pot green synthesis of Ag, Au and Au–Ag alloy nanoparticles using isonicotinic acid hydrazide and starch. Carbohydrate Polymers, 111, 734–743. doi:10.1016/j.carbpol.2014.04.105
Martin, J. D. (2020). Nuclear Magnetic Resonance Spectroscopy. (C. C. M. Mody, Ed.). Between Making and Knowing, 561–569. doi:10.1142/9789811207631_0049
Moorman, V.R., Brayton, A.M. (2021). Identification of individual components of a commercial wheat germ acid phosphatase preparation. PLOS ONE, 16(3). doi:10.1371/journal.pone.0248717
Richardson, A.E. and Simpson, R.J. (2011). Soil Microorganisms Mediating Phosphorus Availability Update on Microbial Phosphorus. Plant Physiology, 156(3), 989–996. doi:10.1104/pp.111.175448
Stücker, A., Podschun, J., Saake, B. and Lehnen, R. (2018). A novel quantitative 31P NMR spectroscopic analysis of hydroxyl groups in lignosulfonic acids. Analytical Methods, 10(28), 3481–3488. doi:10.1039/c8ay01272e
Upadhyay, P., Gupta, M., Simarjeet Kaur Sra, Sharda, R., Sharma, S., Sardana, V., Javed Akhatar and Kaur, G. (2022). Genome wide association studies for acid phosphatase activity at varying phosphorous levels in Brassica juncea L. Frontiers in Plant Science, 13. doi:10.3389/fpls.2022.1056028
Usman, U. (2016). On Consistency and Limitation of Independent t-test Kolmogorov Smirnov Test and Mann Whitney U test. IOSR Journal of Mathematics, 12(4), 22–27. doi:10.9790/5728-1204052227
Van Etten, R.L. and Waymack, P.P. (1991). Substrate specificity and pH dependence of homogeneous wheat germ acid phosphatase. Archives of Biochemistry and Biophysics, 288(2), 634–645. doi:10.1016/0003-9861(91)90246-f
Walker, J. A., Friesen, J. D., Peters, S. J., Jones, M. A., and Friesen, J. A. (2019). Development of a new and reliable assay for choline kinase using 31P NMR. Heliyon, 5(10), e02585. doi:10.1016/j.heliyon.2019.e02585